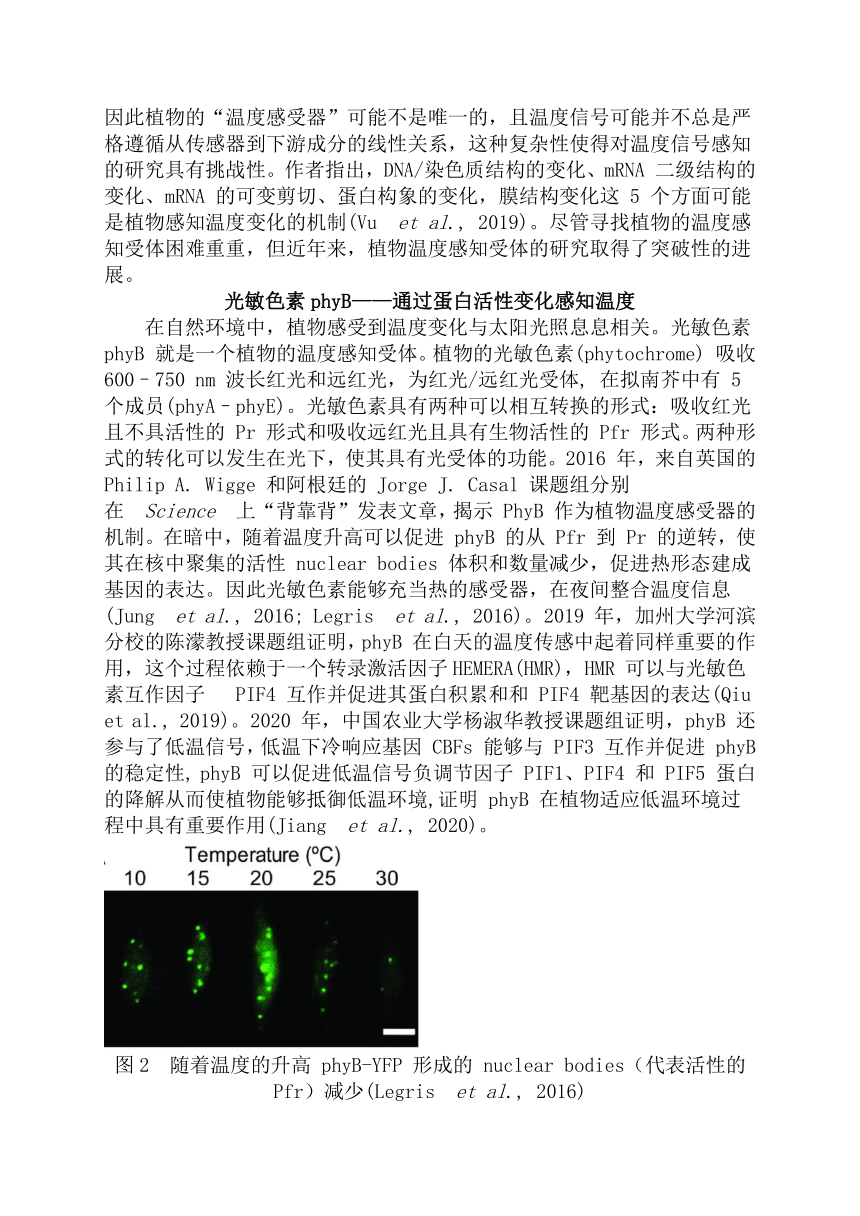
资源简介 植物对温度的感知2019版高中生物学选择性必修一说,温度参与调节植物的生长发育:那么,植物是如何感知温度的呢?地球较为合适的气候为生命提供了繁衍生息的最佳场所,地球的气温会随着太阳辐射、纬度、海陆位置等因素而产生变化。在生物进化的过程中,动、植物分别进化出了不同的机制来感知温度的变化。动物细胞主要通过膜上的 TRP 家族离子通道来作为温度的感知受体。1997 年 David Julius 教授成功筛选并克隆到辣椒素受体 VR1(TRPV1),并证明 TRPV1 不仅能够被辣椒素特异激活,还能被 42 °C 以上高温激活。随后一系列 TRP家族成员被鉴定为温度感受器(ThermoTRPs),这些成员在一定的温度下被激活,共同感知整个生理范围内的温度,其中一些家族成员还可以被薄荷醇、大蒜素等化学配体结合并激活(Castillo et al., 2018)。图1 感知不同温度的 TRPs 通道(Castillo et al., 2018)然而,植物基因组中未发现编码 TRP 家族基因,暗示在植物中存在不同的蛋白或不同于动物细胞的温度感知机制。2019 年 Lam Dai Vu 等人的观点性综述指出,理论上温度可以影响所有生化过程的吉布斯自由能平衡,因此植物的“温度感受器”可能不是唯一的,且温度信号可能并不总是严格遵循从传感器到下游成分的线性关系,这种复杂性使得对温度信号感知的研究具有挑战性。作者指出,DNA/染色质结构的变化、mRNA 二级结构的变化、mRNA 的可变剪切、蛋白构象的变化,膜结构变化这 5 个方面可能是植物感知温度变化的机制(Vu et al., 2019)。尽管寻找植物的温度感知受体困难重重,但近年来,植物温度感知受体的研究取得了突破性的进展。光敏色素phyB——通过蛋白活性变化感知温度在自然环境中,植物感受到温度变化与太阳光照息息相关。光敏色素 phyB 就是一个植物的温度感知受体。植物的光敏色素(phytochrome) 吸收 600–750 nm 波长红光和远红光,为红光/远红光受体, 在拟南芥中有 5 个成员(phyA–phyE)。光敏色素具有两种可以相互转换的形式:吸收红光且不具活性的 Pr 形式和吸收远红光且具有生物活性的 Pfr 形式。两种形式的转化可以发生在光下,使其具有光受体的功能。2016 年,来自英国的 Philip A. Wigge 和阿根廷的 Jorge J. Casal 课题组分别在 Science 上“背靠背”发表文章,揭示 PhyB 作为植物温度感受器的机制。在暗中,随着温度升高可以促进 phyB 的从 Pfr 到 Pr 的逆转,使其在核中聚集的活性 nuclear bodies 体积和数量减少,促进热形态建成基因的表达。因此光敏色素能够充当热的感受器,在夜间整合温度信息(Jung et al., 2016; Legris et al., 2016)。2019 年,加州大学河滨分校的陈濛教授课题组证明,phyB 在白天的温度传感中起着同样重要的作用,这个过程依赖于一个转录激活因子HEMERA(HMR),HMR 可以与光敏色素互作因子 PIF4 互作并促进其蛋白积累和和 PIF4 靶基因的表达(Qiu et al., 2019)。2020 年,中国农业大学杨淑华教授课题组证明,phyB 还参与了低温信号,低温下冷响应基因 CBFs 能够与 PIF3 互作并促进 phyB 的稳定性, phyB 可以促进低温信号负调节因子 PIF1、PIF4 和 PIF5 蛋白的降解从而使植物能够抵御低温环境,证明 phyB 在植物适应低温环境过程中具有重要作用(Jiang et al., 2020)。图2 随着温度的升高 phyB-YFP 形成的 nuclear bodies(代表活性的 Pfr)减少(Legris et al., 2016)ELF3——通过相分离感知高温温度变化是植物生物钟感知的一部分。拟南芥生物钟晚间转录抑制复合物(evening complex) 不仅参与了生物钟的调控,还参与了温度响应。这一复合物包含 ELF3、ELF4 和 LUX 三个蛋白,可以在晚间抑制 PIF4 基因的表达。高温能够解除复合物对于 PIF4 基因的抑制,然而其机制尚不清楚。2019 年英国剑桥大学 Philip A. Wigge 课题组在 Nature 发表文章揭示高温通过促进 ELF3 的相分离调控植物的热响应。ELF3 含有多聚谷氨酰胺的重复序列(polyQ),为预测的朊病毒类似结构域(PrD),polyQ 的长度影响 ELF3 的活性。进一步发现,ELF3 在高温下会聚集形成点状,并且这种聚集的形成依赖于 polyQ,随着 polyQ 长度的增加形成亮点的趋势越大。体外实验进一步证明,ELF3 的 PrD 结构域可以随着温度升高可逆地形成液滴,说明温度可以促进 ELF3 的相分离。本文揭示了一种新的热感性机制:温度通过调控 ELF3 的相变在活动和非活动状态之间快速转变。有意思的是,各种拟南芥的生态型和生活在不同地方的植物具有不同长度 polyQ 结构域,这与不同植物的耐热性相关。(Jung et al., 2020)图3 植物通过 ELF3 的相分离响应高温促使拟南芥开花(Alberti, 2020)COLD1——植物低温感受器2015年中科院植物研究所种康院士课题组报道发现水稻 COLD1 为植物低温感受器。COLD1 编码一种定位于质膜和内质网的跨膜蛋白,其突变导致水稻对冷害敏感。研究发现,COLD1 能与植物中 G 蛋白异源三聚体的 α 亚基RGA1 发生互作,促进 G 蛋白 GTP 酶活性,并激活响应低温的 Ca2+ 通道。COLD1 基因第四个外显子的单个核苷酸的突变影响籼稻、粳稻的耐寒性差异,同时还揭示了遗传进化过程中,来源于中国野生稻品种的 O. refipogon的 CLOD1 基因赋予粳稻的耐寒性(Ma et al., 2015)。近期,种康院士课题组进一步揭示 COLD1 通过调控下游维生素 E-维生素 K1 的代谢调控水稻的低温耐受性。然而目前对 COLD1 感知低温的分子机制及受 COLD1 激活的 Ca2+ 通道仍不清楚(Luo et al., 2021)。未来,需要通过结构生物学等手段进一步阐释低温激活 COLD1 的原理和构象变化。图4 粳稻在驯化过程中通过 COLD1 基因位点的选择来适应低温(Li and Lin, 2016)植物固着生长的特点迫使植物进化出更加复杂的温度感应机制以应对复杂的环境变化。对植物温度感知的研究将有助于我们认识植物适应环境的机制,为作物品种改良提供重要的理论基础。例、下列关于植物感温性的描述,正确的是( )A.感温性是植物对环境温度规律性变化的适应B.在一定温度范围内,随温度升高,植物生长发育速度加快C.有些植物或品种在较高温度刺激下发育加快,即感温性较弱D.晚稻的感温性强于中稻,中稻强于早稻答案:ABD 展开更多...... 收起↑ 资源预览
 资源预览
资源预览

 资源预览
资源预览